As the
central nervous system develops, the axons of each retina’s
ganglion cells form the optic nerve. When the
growth cones of these axons reach the optic
chiasm, they must “decide” whether
or not to cross over to the other side of the brain. As the
growth cones of these axons approach this decision point,
they slow their advance and assume a more complex form.
The same phenomenon is observed in
the development of the peripheral nervous system, where
the growth cones of the motor neurons start to engage in
more “seeking” behaviour when they enter the
newly forming muscles of the arms and legs.
THE GROWTH CONE
When
the neuroblasts have
completed their
migration, or even while they are still making it,
they send out extensions called neurites that
grow from their tips. One of these neurites, which will
become the axon, will have to grow a long distance before
reaching its target. Its elongation will be made possible
by a structure at its tip, called the growth
cone.
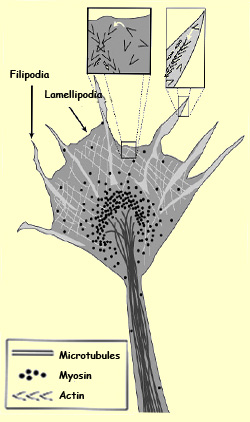
The growth cone of an axon (or of a dendrite) is composed
of flat, fanlike membranes, the lamellipodia,
from which fine tubes called filipodia protrude.
These filipodia extend and retract constantly to explore
their surroundings. When a filipod, instead of retracting,
attaches itself to the substrate, it makes the growth cone
advance in that direction.
When axons are growing along a pathway already established
by other axons, their growth cones are rather simple
in form. But if an axon starts to open up a new pathway,
or if it arrives
at an intersection where it has to pick a direction,
its growth cone becomes spectacularly complex. It flattens
and sends out numerous filipodia to actively search
for the signals that can guide it (see sidebar).
Actin is represented in
light grey on the main drawing, and by the black V
shapes ( >>>>>) in the enlargements.
The white arrows in the enlargements represent the
polymerization of the actin.
The growth cone responds to various
molecular signals that show it what path to follow and, at the
end of its journey, help the axon to form
proper synaptic connections. In this process, as in neurotransmission,
the affinity between these guidance molecules and their receptors
on the growth cone membrane plays a vital role. The stimulation of
these receptors causes the activation of second messengers that trigger
the intracellular events responsible for determining the direction
of this growth. These events are thought to involve reorganizations
of elements in the axon’s cytoskeleton.
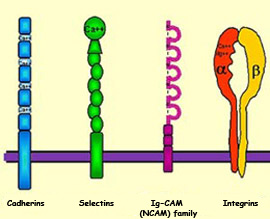
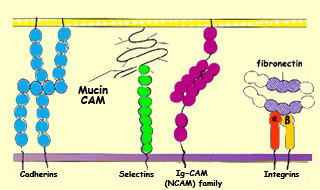
One of the characteristics
of almost all CAMs is that their extracellular portions
contain repeated patterns of amino acids. CAMs are divided
into two families, however, according to whether they need
calcium to adhere to cells: cadherins do
need calcium (they are “calcium-dependent”),
whereas neural cell adhesion molecules (NCAMs)
do not (they are “calcium-independent”).
CAMs are transmembrane proteins that protrude from the
surface of cells in the growth cones’ environment.
CAMs can thus interact with CAM-specific receptors on
the growth cones.
When a CAM and its receptor recognize each other, a biochemical
cascade of second messengers is triggered within the growth
cone. This cascade results in the activation of enzymes (kinases,
phosphatases, proteases, etc.) whose effects will contribute
to the elongation of the axon.
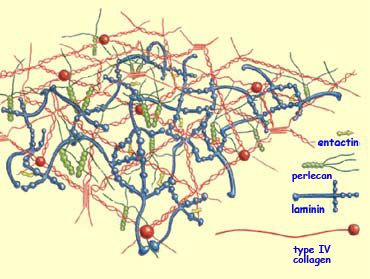
Some receptors on growth cones are also sensitive to proteins
that are not located on cell membranes but are instead
distributed in the extracellular matrix—an agglomeration
of substances produced by cells but not directly attached
to them.
The best known of these adhesion molecules in the extracellular
matrix are the laminins, the collagens, and fibronectin.
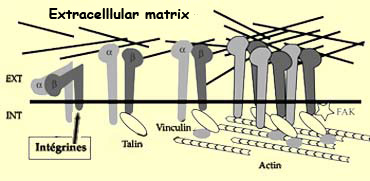
These molecules are specifically recognized by a class of
growth cone receptors called integrins.
In addition to surface proteins, another important class of molecules
can influence the direction in which the axon elongates. These
molecules, called chemotropic factors, are secreted
by the target cells in very small amounts and diffuse into the
surrounding extracellular environment. They can be either
chemoattractive and attract the axon or chemorepellent and repel
it.
Note that these chemotropic factors are not the same thing as another class of
diffusible molecules called trophic
factors. The function of trophic factors, of which nerve growth
factor (NGF) is one, is to keep the neuron alive and to facilitate the growth
of its axons and dendrites.
BDNF is especially
important for the survival of the neurons
of the visual cortex. Most of the receptors
to which neurotrophins bind, known as trk receptors, are
kinase proteins that phosphorylate tyrosin residues found
in other proteins that form their substrate. In other words,
they add phosphorus atoms to the amino acid tyrosin in
certain proteins to modify their form and hence their function.
In the case of the development of the visual cortex, this
phosphorylation ultimately has an effect on gene expression.
Neurotrophins, and
in particular BDNF, which is very widely expressed in the
central nervous system, appear to play a significant role
in synaptic plasticity:
the morphological and physiological changes that synapses
undergo in response to changes in neuronal activity. We
know, for example, that the synthesis and release of BDNF
molecules by the neurons of the central nervous system
are controlled by neuronal activity, which enables this
neurotrophin to modulate the GABAergic and glutamatergic
transmissions of certain brain structures such as the hippocampus and
the visual
cortex.
TROPHIC
FACTORS AND NEURONAL DEATH
Trophic factors (also called growth factors)
contribute to the development and maintenance of the body’s
networks of neurons, but are not the same as axon
guidance molecules. Trophic factors are another class of molecules
that are secreted by target cells. The role of these molecules
is not to help the axon orient itself, but rather to ensure its
survival once it has formed certain functional synaptic connections.
Trophic factors are secreted in limited amounts by the
target cells, so that only a subset of the neurons that
innervate them receive enough to survive. In other words,
every neuron needs a certain minimum amount of trophic
factor to survive, and all indications are that the neurons
compete for the trophic factors available. The neurons
that fail to get enough simply disappear, by apoptosis,
the body’s programmed process of cell death (for more on
apoptosis, follow the Tool Module link to the left).
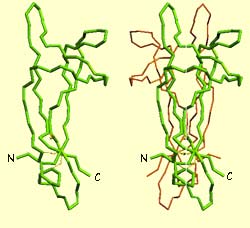
The first trophic factor ever discovered was nerve growth factor (NGF).
NGF was identified as a protein composed of three sub-units,
of which one in particular is truly indispensable for the neurons’ survival. This sub-unit,
designated
ß, is itself composed of two identical molecules of
118 amino
acids).
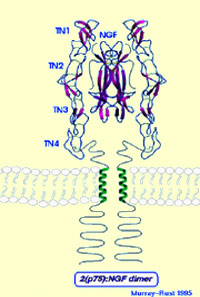
The two green diagrams on the left represent
the molecular structure of NGF, which consists of two symmetrical
components connected to each other along their longitudinal axis.
The blue diagram on the right shows an NGF molecule bound to the
centre of an NGF receptor.
Though NGF is the prototypical trophic factor and probably the
one that has been studied the most, it affects only certain categories
of peripheral neurons. Since the mid-1980s, other trophic factors
related to NGF have been identified in many studies. This family
of molecules is now known as the neurotrophins.
In addition to NGF, its includes three other molecules that have
been well characterized: brain-derived neurotrophic factor (BDNF),
neurotrophin-3 (NT-3), and neurotrophin-4/5 (NT-4/5).
Indeed, experiments have shown that the growth
of neurites can be controlled locally by growth factors, without
necessarily involving the enzymatic mechanisms inside the cell
body. Consequently, some of a neuron’s axons or dendrites
may be extending at the same time as others are retracting, as
is in fact seen during the formation of synapses.
The affinity between
an axon and its target cell is something like the colour
coding used in multi-wire electrical cables so that the
right wires can be connected to each other.
But whereas the coded colours in electrical wiring are designed
to be mutually exclusive, research has shown that the affinity
between neurons and their target cells is not highly selective.
Certain axons do show a preference for certain target cells
but can also establish synaptic connections with other neurons.
Associations between neurons and
their target cells thus take place along a continuum
of preferences. At one end of this continuum, axons are
absolutely unable to make connections with glial cells,
for example, while at the other end, axons can make connections
to any cell at all within a given population.
When a neuromuscular
junction is forming between the end plate
of a motor neuron and a muscle fibre, even the biological
properties of the nicotinic acetylcholine receptors
on that fibre change. Their replacement rate decreases,
their ability to pass ions increases, and the combination
of sub-units that compose them is altered. All of these
changes contribute to the fine-tuning of the neuromuscular
junction.
FORMATION AND SELECTIVE STABILIZATION
OF SYNAPSES
When a growth cone
first makes contact with its target cell, the synapse does
not become functional immediately. The formation of a synapse
is a gradual process. This process has been studied extensively
in one particular class of synapses: neuromuscular junctions.
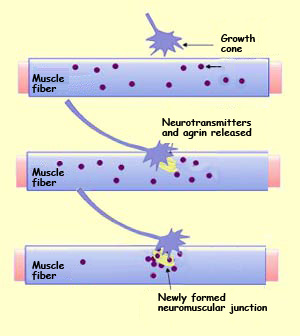
We know, for example, that the growth cone of a motor
neuron secretes acetylcholine spontaneously, before even reaching the muscle
fibre with which it will form a neuromuscular junction. We also know that initially,
the nicotinic
acetylcholine receptors on this muscle fibre’s membrane are distributed
uniformly. But shortly after the motor neuron’s axon
makes contact with the muscle fibre, nicotinic receptors accumulate
rapidly on this fibre at the site where the neuromuscular junction
will form, while the population of such receptors away from
this site declines drastically.
In addition to redistributing its receptors, the postsynaptic
cell forms new receptors in its membrane at this site. It
follows logically that messenger
RNA for these receptors is being synthesized in the cell
nucleus closest to the developing neuromuscular junction.
Initially, the development of neural pathways is controlled
by genetically programmed mechanisms. But the neural circuits
formed by these intrinsic mechanisms are still only crudely laid
out and contain myriad extremely redundant synapses. To reduce
the number of synapses and refine these circuits, a selection
process is required.
This selection process depends on the activity of the
neurons; it is thus through the individual’s sensorimotor
experience that the initial circuitry will be tested
and the fine structure of the neural networks will be
adjusted. But how exactly does the activity of the neurons
as they respond to their environment affect the development
of these circuits?
To answer this question, we must turn to Hebb’s
postulate. Originally formulated to explain the cellular bases for learning
and memory, this postulate also applies to the major synaptic changes that
occur during the development of the nervous system. According to Hebb, the correlated
activity of two neurons causes a synapse to be strengthened. As applied to development,
this postulate means that if two neurons that are connected to the same target
cell transmit co-ordinated signals, both of their synapses will be reinforced.
Conversely, if their signals are out of phase, these synapses will be weakened.
Consequently, in the course of development, those synaptic endings whose activity
is only rarely correlated with that of the postsynaptic neuron will gradually
weaken until they disappear completely. This phenomenon was given the name selective
stabilization of synapses by Changeux and Danchin (1976), who showed
that only those synaptic connections that are incorporated into a functional
neural circuit will survive.
In contrast to neuronal
death, which serves to adjust the number of neurons to
the number of target cells, the elimination of synapses serves
to make the pattern of innervation more precise.
Throughout our lives,
but especially during infancy, our synaptic connections
are shaped by our sensory experience. Neurons can increase
the efficiency of these connections through the process
of long-term
potentiation (LTP), or they can decrease it
through the process of long-term
depression (LTD). Both of these processes contribute
to the fine-tuning of our neuronal connections, but LTD
seems to play an especially important role in the selective
elimination of synapses that characterizes certain critical
periods of human development.
LTD leads to a reduction in the number of postsynaptic receptors,
which would reduce the activity of the synapses concerned
and could lead to their gradual elimination as observed during
these critical periods.