|
|
| Funding for this site is provided by readers like you. | |
|
|
|
|
|
|||||
|
|
|||||||
|
|
|
|
|
|
A
Monthly Podcast On Cognitive Science Brain Rhythms: The Oscillations That Bind The “Coming Out” of the Electrical Synapse
|
|
The binding problem asks the question: how can we have a conscious, coherent, unified perception of an object, given that its various attributes are processed in different parts of the brain? One approach to an answer would be to say that the various signals from all these different parts of the brain converge on a group of cells or even a single cell that represents this conscious perception. Indeed, though the brain has a preponderance of parallel circuits, it does display a certain form of convergence as one moves farther from the primary areas of the cortex and closer to the “associative”ones. Many such areas of convergence have been identified in the frontal cortex, the anterior temporal cortex, and the inferior parietal cortex. But neuroscientists agree that these are not sites where mental representations are stored. At most, they may contain certain “codes”that can reconstruct the fragments of activations distributed elsewhere in the sensorimotor areas of the cortex. Likewise, it is true that there are certain neurons located at the top of the hierarchy in the visual cortex that respond specifically to faces, and even to faces seen from a certain angle. These neurons definitely help us to recognize faces, because when these neurons are destroyed by a stroke, the stroke victim develops prosopagnosia, the inability to recognize faces, even of immediate family. But apart from a few special cases such as recognizing faces, which has always been of tremendous adaptive importance, convergence is too flimsy, inefficient, and ultimately ineffective a mechanism for unifying perceptions in the brain, and it does not seem to have been selected by evolution for this purpose (follow the link to the left). So if there is no single place where all of the information about an object converges to become conscious, is there perhaps a single time when it does? That is the other major approach to solving the binding problem, and it seems the more promising. In this approach, broadly speaking, neurons that are active at the same time are believed to be “perceiving the same thing”. In technical terms, this approach is based on the temporal synchronization of neuronal activity.
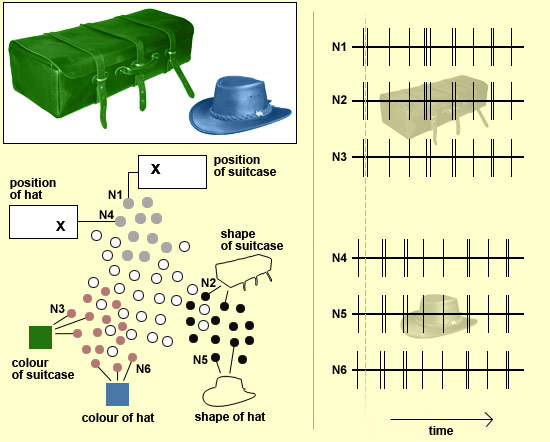
To return to the example of the green suitcase next to the blue hat, each of these two objects will be represented by a huge assembly of neurons in the brain. Each of these assemblies will include neurons that can detect various attributes of each object, such as colour or movement, or the orientation of its lines and contours, and so on. And thus, according to this hypothesis, it is through the synchronization of the various neurons coding for the various attributes of the suitcase (N1, N2, and N3 in the figure below) that the brain obtains a coherent, unified image of this object. As for the neurons that code for the various attributes of the hat (N4, N5, and N6), they too fire synchronously to provide a unified image of this object. But as the grey dotted line shows, they do not fire at the same time as the neurons that code for the various attributes of the suitcase. And that may be how we consciously perceive two separate objects standing out against a background that is also a separate object (and that would be represented by a third assembly of neurons), and not simply an undifferentiated amalgam of lines and colours.
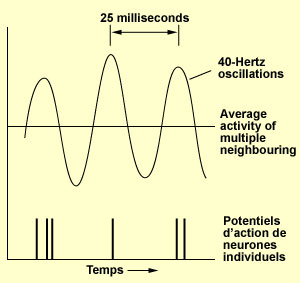
Francis Crick and Christof Koch took the idea of temporal synchronization one step further by proposing that this synchronized activity, when occurring at 35 to 75 Hz (hertz, or cycles per second), may be the neuronal correlate for conscious visual perception. Their idea originated with studies done on the visual cortex of cats in the 1980s. These studies had shown that large numbers of neurons could fire at the same time at a rate ranging from approximately 35 to 75 Hz. (Neuronal activity oscillating at this frequency is commonly referred to as “gamma oscillations” or simply “40-Hz oscillations”.) Many subsequent studies both in animals and in humans showed that this high oscillation frequency of neuronal activity is closely related to the integration of perceptions, the construction of coherent representations, and to some processes of selective attention. Crick and Koch therefore developed a theory according to which the key to conscious perception lies not solely in the synchronization of neuronal activity, but in the synchronization of neuronal activity oscillating at frequencies in the range of 35 to 75 Hz. In summary, according to this hypothesis:
Crick and Koch also believe that these transitory neuronal assemblies oscillating at about 40 Hz do not form only in the visual cortex but may also recruit neurons throughout the cortex. In this case, the colour and shape of an object would not be the only characteristics associated with it. There would also be all sorts of other characteristics, such as odour, taste, emotional associations, and so on, all of them thus helping to form a complete conscious representation of the object observed. Here then is an elegant mechanism by which the brain might distinguish, among all the representations that it has bound, those that are conscious from those that are not. Crick and Koch had first proposed, in the late 1980s, that synchronization of the oscillations around 40 Hz was a sufficient mechanism to ensure the emergence of a conscious perception. Shortly after the year 2000, they altered their position somewhat, stating that conscious phenomena seem to arise from a competition among various “coalitions” of neurons (see box below) in which the winning coalitions determine the content of the conscious mind at any given time. And what these authors now consider a more plausible role for the synchronization of the 40 Hz oscillations is that it might act as the mechanism by which this competition is resolved, by favouring the selection of a particular neuronal assembly. This idea of neuronal assemblies that are competing at all times is also consistent with the more recent hypothesis of the “dynamic nucleus”. Engel and Wolf also believe in synchronization, though they consider it necessary but not sufficient to generate consciousness. In their view, the information should also enter into a form of short-term memory, a suggestion that here refers to something like a global workspace. In short, though the the hypothesis of the
synchronization of oscillations at around 40 Hz undeniably contributes
something to our understanding of consciousness, it is not the final
word on this subject. It has, for example, undergone further interesting
developments, particularly as regards the mechanism
by which the brain selects the representations that will become
conscious from among all of its unconscious
representations.
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

| |
|
|
|
|
|
|
|
|