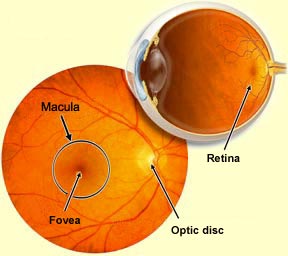
The blood vessels on the retina's surface
can be seen with an ophthalmoscope (the instrument that physicians
use to examine the back of the eye through the pupil). These
blood vessels enter the eye through a colourless region of
the retina, known as the optic disc. The
optic disc is also the head of the optic nerve; it is here
that the axons of the retina's ganglion cells converge and exit the
eye.
Because there are no photoreceptors in the optic disc, this
part of the retina is insensitive to light, just as it is
at the location where the largest blood vessels pass. For
this reason, the optic disc is also known as the blind
spot. But you do not experience any interruption
in your field of vision at your eyes' blind spots, because
the brain has a way of "filling in" your visual perceptions at these locations
(see the first Experiment module link to the left).
At the centre of each retina there is
also a darker area called the macula that is practically
devoid of blood vessels, in order to optimize central vision (as
opposed to peripheral vision). At the centre of the macula, a small
depression of about 2 mm in diameter forms the fovea—the
part of the retina that is composed exclusively of cones and where visual acuity is greatest.
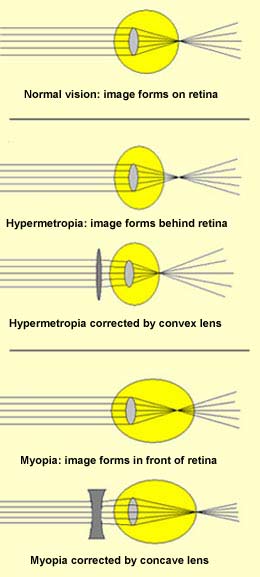
When light rays do not converge on the retina precisely,
several different types of vision defects may occur.
For example, if the eyeball is too short from front to
back, the rays converge at a point beyond the retina. This
defect is called hypermetropia, or far-sightedness.
It can be corrected by eyeglasses with convex lenses, which
increase the rays' convergence and thus pull the focus
back onto the retina.
If the eyeball is too long from front to back, then the
rays converge in front of the retina. This condition is
called myopia, or near-sightedness. It
can be corrected by eyeglasses with concave lenses.
Another vision defect, presbyopia, is
due to hardening of the lens, associated with aging. As
the lens hardens, it becomes less elastic, so that it can
no longer assume a sufficiently convex shape during accommodation or a sufficiently flat
one during relaxation. The eyeglasses prescribed for this
condition contain bifocal lenses, where the top half is
concave for distance vision, and the bottom half is convex
for close-up vision.
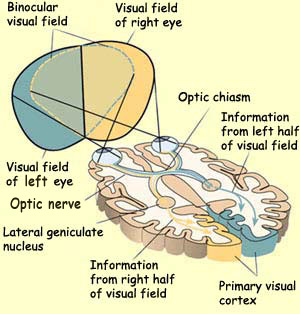
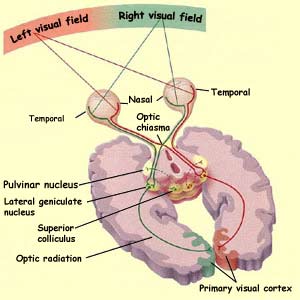
The visual field or
receptive field of an eye is the area in space that is
imprinted on the retina when the eye is focused on a distant
point. The visual fields of the two eyes overlap to a large
extent, but the right eye's field extends farther to the
right, and the left eye's extends farther to the left.
The figure above also shows how the right eye's visual
field (in yellow) is analyzed in the left visual cortex, and vice versa. The central
area where the two eyes' visual fields overlap is the binocular
visual field.
The numerous connections among the
various areas of the brain that are involved in processing
visual information (such as the visual cortex, basal
ganglia, pons, cerebellum,
and oculomotor nuclei) are generally reciprocal. This reciprocity
creates feedback loops which vividly demonstrate just how
much the visual system is intrinsically interlinked with
the operation of the nervous system as a whole.
The evolutionary history of the human brain sheds
much light on the reason for all these neuronal feedback
loops in our visual system. In the reptilian brain, for example, vision is tightly
linked with reflex defence responses. A deafening noise,
a new tactile sensation, or an object approaching rapidly
from the edge of the organism's field of vision will make
it turn its head quickly toward the new stimulus so that
the eyes can assess how much danger it actually represents.
Even though the visual system of humans and other primates
has become far more sophisticated to let us acquire a conscious,
detailed vision of the world around us, these old circuits
are still useful, have been preserved by evolution, and are
still at work in the human brain.
THE TARGETS
OF THE OPTIC NERVE
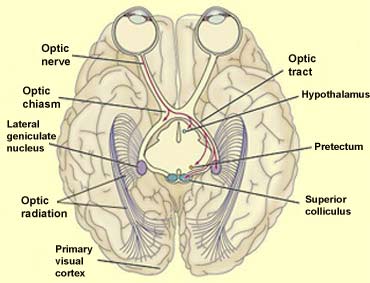
The axons of some of the ganglion cells of the retina diverge from
the optic tract to project to structures other than the lateral geniculate nucleus, which is the main
relay between the retina and the visual cortex.
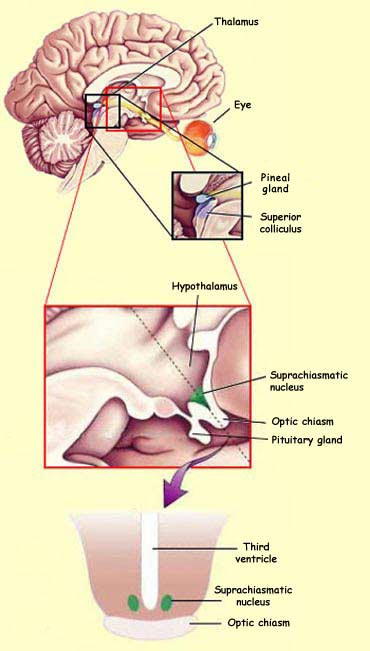
One of these structures
is the hypothalamus, and more specifically
its suprachiasmatic nucleus, which receives
a certain number of connections from retinal axons. The
suprachiasmatic nucleus is considered the primary site
of the body's internal biological clock. The visual signals
that it receives from the retinal axons keep it continuously
informed about the darkness or lightness of the environment,
thus enabling it to synchronize a wide range of biological
rhythms, including sleep and wakefulness, that are linked
to the cycle of day and night.
Axons from some other retinal ganglion cells project to
the pretectum, a part of the midbrain
that controls the opening of the pupil and certain eye
movements.
Lastly, about 10% of the axons emerging from the retina
project to a part of the tectum (roof) of the midbrain
called the superior colliculus. This pathway
is relatively large. It comprises about 150 000 axons,
equivalent to the total number of ganglion cells in the
retina of a cat! In fact, the superior colliculus corresponds
to the optical tectum in all non-mammalian vertebrates,
in which this retinotectal projection is the main efferent
pathway from the retina.
Because of the way that receptive fields overlap one another
in the retina, a beam of light shined on the retina
activates a large population of neurons in the superior
colliculus. These neurons trigger movements of the eyes
and head, via the motor neurons of the brainstem, to try
to bring the image of the light beam onto the fovea. Thus the retinotectal pathway
is involved in orienting the eye toward a stimulus that
initially appears in its peripheral field of vision.
Like the lateral geniculate nucleus, the superior colliculus
also receives connections from the primary visual cortex.
The neurons of the superior colliculus in turn project
their axons to subcortical structures such as the reticular
formation, the inferior colliculus, and the spinal cord.
The neurons of the superior colliculus also influence two
structures that are involved in vision: the lateral geniculate
nucleus and the pulvinar.
The pulvinar is a nucleus in the posterior
portion of the thalamus. It receives afferences directly
from the optic tract (via collateral axons) as well as
by way of the LGN. Like many other thalamic nuclei, the
pulvinar was long regarded as a relatively passive relay
for information en route to the cortex, where the real
information processing was assumed to take place. But
this view of the pulvinar has been altered radically
by the accumulation of data showing that its neurons
display sophisticated visual responses of which formerly
only the cortex was thought to be capable.
Now the pulvinar is instead believed
to be an image-interpreting centre that plays an important role
in visual attention and motion perception. For example, the pulvinar
is thought to help maintain the stability of the body's visual
environment by compensating for the effects that body movements
have on the position of images on the retina. Hence it is no surprise
that the neurons of the pulvinar have been found to project to
the secondary
visual areas involved in detecting motion.
Visual agnosia,
which often occurs following bilateral occipito-temporal
lesions, makes people incapable of discriminating the shapes
of objects, though these people may sometimes retain good
abilities to discriminate colours and textures. People
with visual agnosia perform very poorly when asked to recognize
the shapes of arbitrary geometric objects, letters of the
alphabet, and black and white drawings, yet have no trouble
in moving their hand toward an object, turning their wrists
to insert an object in a slot, or arranging their fingers
in the right way to pick up an object. Visual agnosia syndrome
thus impairs conscious visual awareness but leaves the
ability to manipulate objects intact.
In the opposite syndrome, optical ataxia,
people with damage to their parietal lobe can recognize objects
but cannot pick them up and use them appropriately. Observations
of such individuals have helped researchers to uncover the
differing roles of the ventral and dorsal visual pathways.
In 1983, Joseph Zihl and his collaborators
published an article in Munich about a 43-year-old woman
who had become totally incapable of perceiving movement,
following a stroke that had damaged both sides of area
V5, the part of the extrastriate cortex that is involved
in recognizing movement. This patient was thus suffering
from the strange syndrome known as akinetopsia,
or motion blindness. For several seconds
at a time, she would see nothing but a still image and
have no conscious awareness of any movements in her environment.
Crossing the street, for example, was very dangerous for
her, because a car that she had last seen "stopped"
a good distance away might suddenly appear right on top of
her after she had begun to cross. Pouring a glass of water
could be almost as big a problem. Since she saw the water
as frozen in space rather than flowing, she could not tell
that she had overfilled the glass until she suddenly saw
a puddle of water on the table.
THE VARIOUS
VISUAL CORTEXES
Following the groundbreaking studies
published by Leslie Ungerleider and Mortimer Mishkin in 1982, scientists
distinguished two major pathways for the cortical processing of
visual information: the ventral visual pathway, for identifying
objects, and the dorsal visual pathway, for determining their position
in space. Various subsequent studies, however, have raised some
questions about this dichotomy. Some of these studies have involved
making selective lesions in each of these pathways in monkeys.
Others have involved observing humans who had suffered brain injuries
that affected only one of these pathways (see sidebars).
Today it is believed that the main function of the dorsal visual pathway is
to guide in real time the actions that we direct at objects
in the visual world. Most of the processing done by this pathway
is believed to be unconscious. The dorsal pathway could thus
be described as an "action pathway", because
by integrating the spatial relationships between our bodies
and our environment, it lets us interact with this environment
effectively. The ventral visual pathway, on the other hand,
seems to be involved in forming conscious representations of the identity of
objects. Thus, in addition to the functional dichotomy between the dorsal and
visual pathways, there would appear to be another dichotomy, between unconscious
and conscious vision.
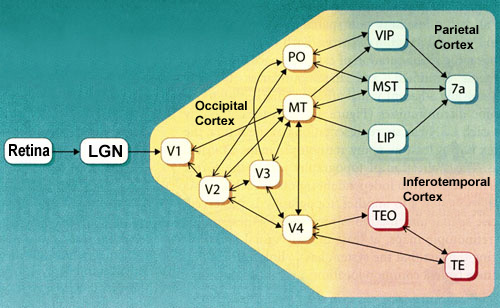
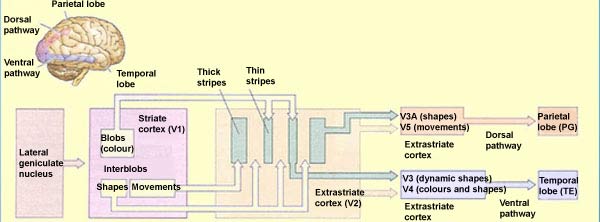
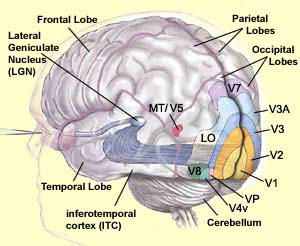
The dorsal pathway comprises several cortical
areas, including the medial temporal area (MT or V5), the medial
superior temporal area (MST), and the ventral and lateral intraparietal
areas (VIP and LIP).
Area V5 (or MT) seems to contribute significantly
to the perception of movement. This area receives projections
from V2 and V3. It also receives projections from layer IV B in the primary visual area (V1)—a
layer that, interestingly enough, is part of the magnocellular
channel involved in analyzing the movement of objects. This channel
also maintains its specificity for movement in area V2, where
it is concentrated in the thick stripes that contain large amounts
of cytochrome oxydase.
It has therefore been suggested that the
segregation between the magnocellular and parvocellular signals persists
up to the highest levels of visual analysis. The marked functional
difference between the ventral and dorsal pathways might also
be attributed to a preferential contribution from the P-IB channel
in the former case and from the M channel in the latter.
For the cells
in area MT, the movement of an object seems more important
than its nature—so much more, that this area of the
cortex is organized into columns that code for the orientation
of the movement, just like the line-orientation columns in V1.
Some cells in area MT even seem to respond not to the actual
direction of movement, but rather the perceived direction.
For example, two groups of illuminated lines, each moving
at 45 degrees to either side of the vertical, will give
the impression of a vertical movement as they intersect.
In area V1, the cells that have a preference for 45-degree
angles will respond best to this kind of stimulus. But
in area MT, many cells that normally show a selectivity
for vertical directions will respond convincingly to two
stimuli moving at 45 degrees—in other words, to the
apparent direction of movement.
Beyond area MT, there are other areas
involved in analyzing movement, such as area MST.
The cells in this area are sensitive not only to linear motion,
like the cells in area MT, but also to radial motion (to or away
from a point) and circular motion (clockwise or counter-clockwise).
They are also selectively activated by complex configurations of
movements, such as occur in your surroundings as you walk through
them.
Certain neurons in the superior temporal polysensory area (STP) even respond
selectively to biological movements that may have proven essential for individual
survival, such as by recognizing a member of the same species from the way
that he or she walks.
Your brain may use this large volume of movement-related data gathered by the
dorsal pathway for many purposes: to extract relevant information about the objects
flowing across your field of vision as you move, in order to guide your movements;
to orient your eye movements; or to identify moving objects around you that have
implications for your survival.
In addition to the serial path
that visual signals follow up through each of the hierarchical
levels of the visual system, there are also numerous
channels that process information in parallel, thus forming
a network of highly complex circuits.
This complexity is at least partly attributable to the
numerous feedback loops that each of these areas forms
by returning connections to the areas that send connections
to it. Another aspect of this complexity comes from the
projections to subcortical structures such as the lateral geniculate nucleus and the superior colliculus.