The light that enters the eye must pass through all
the layers of the retina before reaching the
outer segments of the photoreceptors. The inner segments of
the cones, located just in front of the photosensitive pigments
on the light rays' path, seem to act like optic fibres, guiding
the light rays to the outer segments. This would explain why
the cones are far more sensitive to light rays from the centre
of the eye. These rays are better aligned with the cones'
inner segments and hence pass through them more easily than
rays that strike them obliquely. The cones thus take advantage
of the better optics at the centre of the eye.
In contrast, for the rods, because of their essential role
in low light conditions, every photon is important, and they
cannot afford to lose any. That is why the rods do not have
a selective-direction system like that found in the inner
segments of the cones.
New discs are constantly being produced at the base of the outer segments of the rods and cones, so these segments are constantly growing longer. That is why the pigment epithelium
that is in contact with the old discs at the tip of the outer segment phagocytoses ("eats") and destroys them.
PHOTORECEPTORS
The transduction (conversion)
of light signals into nerve impulses is accomplished by some
125 million photoreceptors located in the deepest part of the retina.
This task is divided between two
types of photoreceptors that are very different from each other.
The 120 million receptors called rods let
you see shades of grey in low light conditions (whence the
saying "All
cats are grey in the dark"). The other 5 million receptors,
called cones, are smaller and wider, and sensitive
to colour in bright light conditions.
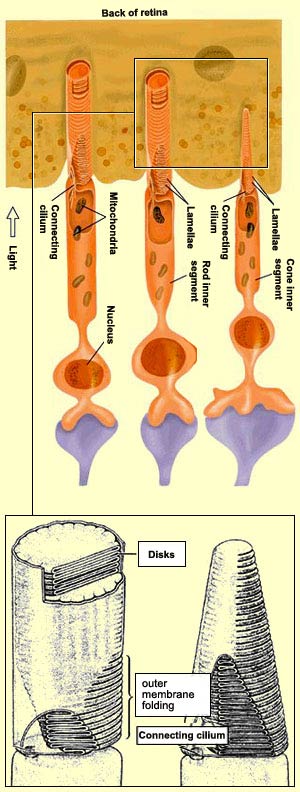
The outer segments
of the rods are cylindrical, while those of the cones are cone-shaped.
But shape is not the only feature that distinguishes the two.
They also differ in the number and arrangement of the discs
formed by the folding of their membranes. In the rods, there
is a stack of about 900 of these discs, which become completely
detached from the membrane and float freely inside it. In the
cones, there are far fewer discs, and instead of becoming detached
from the outer segment membrane, they remain attached to it.
These photoreceptors are actually nothing more than highly
specialized cilial cells whose outer and inner segments are
joined by a connecting cilium. The inner segment of each photoreceptor
contains the cell's nucleus and organelles such as mitochondria
and Golgi bodies that are essential for the functioning of
any cell.
In the inner segments, as in the outer ones, there are some
notable anatomical differences between rods and cones (see
sidebar).
The
distribution of rods and cones varies from
one point to another on the retina's surface. There
are very few cones around the periphery, where
rods predominate. In contrast, in the central region
of the retina, called the fovea,
there are no rods at all. That is why you turn
your eyes to make an object that you want to look
at fall within this area of greater acuity within
your field of vision.
Lastly, the most important functional distinction between rods
and cones, the one that makes cones sensitive to colours whereas
rods are not, comes from their differing photopigments. While all
rods have the same kind of photopigment, called rhodopsin, the
outer segments of cones contain one of three different opsins that
have absorption peaks in the short, medium, and long wavelengths
of light, respectively. These three pigments, with their differing
spectral sensitivities, are the basis for human
colour vision.
The eye's sensitivity
to light is not constant. Instead, it adjusts to light
levels in various ways. Dark adaptation occurs,
for example, when you enter a darkened movie theatre
when the film has already started. At first you feel as
if you can't see anything. But very quickly, your irises
open to let more light reach your retinas.
A slower adaptation also takes place, involving the photosensitive
pigment in the rods, rhodopsin.
Over the first 20 to 25 minutes that you spend in a dark
environment, because your rods' stores of rhodopsin are
no longer being bleached by the light, they regenerate
more readily, causing the rods' sensitivity to light to
increase about a million-fold!
When you come back out of the theatre into the daylight after
your eyes have adapted to the darkness, you are temporarily
blinded, until they have completed the reverse process, light adaptation. In
the first step of this process, your irises close rapidly
to reduce the amount of light entering the eyes. Next, the
other biological changes that occurred in dark adaptation
are reversed as well, so that after just a few minutes, your
vision has adjusted to the bright light of a sunny day. As
this implies, the cones adapt to light more quickly than
the rods adapt to darkness.
The function of the photoreceptors is
to transduce (convert) light energy into membrane
potential. In many ways, the mechanics of this
process are comparable to those found in synapses
that use metabotropic
receptors to achieve transduction chemically.
When a neurotransmitter binds to a metabotropic
receptor, it activates
G proteins that in turn stimulate various enzymes .
These enzymes alter the intracellular concentration
of a second messenger, which results in a change
in the conductance of certain ion channels and
hence a change in membrane potential.
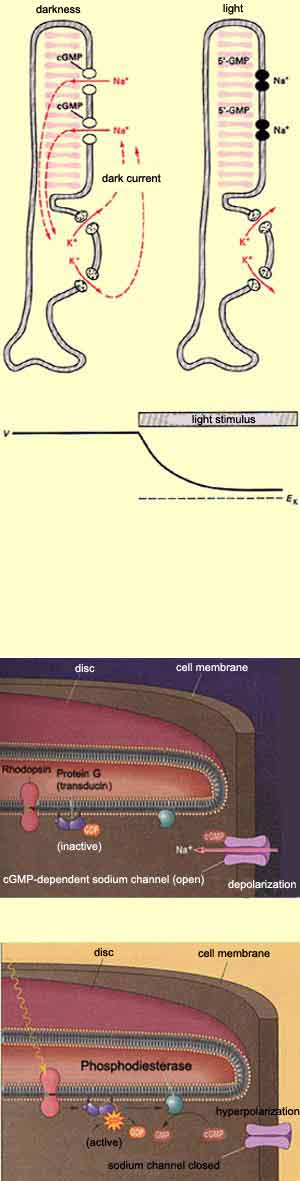
The transduction of light by the photoreceptors
in the retina involves the same basic steps. But
before we describe them, it must be noted that
while the resting potential of most neurons is
usually around
–65 mV, the membrane potential of the
outer segment of rods is about – 30 mV in
the dark. The reason for this depolarization is
that in the absence of light, there is a continuous
flow of sodium ions into each rod's outer
segment through sodium-specific channels in
its membrane. These channels are kept open by the
presence of the second messenger cyclic guanosine
monophosphate (cGMP), which in dark conditions
is produced continuously by the enzyme guanylate
cyclase. This phenomenon is known as the dark
current.
When photons of light strike the light-sensitive
pigment in the rods or cones, it
changes form and thus activates a G protein
called transducin. This transducin in turn activates
the enzyme phosphodiesterase, which metabolizes
cGMP and thus reduces the level of cGMP in the
photoreceptor. This drop in cGMP in turn reduces
the outer segment's sodium conductance and, consequently,
the dark current that is responsible for its unusually
high membrane potential.
The result is thus the contrary of what might be
expected: the presence of light hyperpolarizes
the photoreceptor cell and consequently causes
it to release fewer neurotransmitter molecules
into its synapse with the bipolar
cells.