A large body of evidence indicates that
the dorsolateral prefrontal cortex plays an important role in certain
forms of memory work, in particular those that involve alternating
between two memory tasks and exploring various possibilities before
making a choice.
It seems fairly certain that this area of
the brain holds information required for reasoning processes that
are in progress. But its precise role remains the subject of much
debate. Does this prefrontal area basically coordinate the activities
of slave sub-systems, as in Baddeley’s
model of the phonological loop and the visual/spatial sketchpad?
Or does it actually itself serve as a temporary storage area for
certain types of information, as Goldman-Rakic’s research
tends to indicate? Might the level of abstraction of the task be
the deciding factor, or might the size of the workload determine
whether this area comes into play?
As all these unanswered questions suggest,
the anatomical substrate of working memory is far from being understood
in detail. Moreover, the phenomenon of working memory is made all
the more complex by the fact that it takes place over time.
Source: NIMH Laboratory of Brain and
Cognition. Published in Nature, Vol. 386, April 1997,
p. 610.
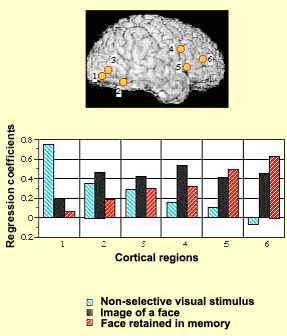
For example, the experimental
results illustrated here show how various areas of the subjects’ brains
alter their activity levels as the subjects are presented
with various visual stimuli. When the subjects are shown
a blurred image, the activity level (represented by the blue
bars in the graph) becomes highest in area 1, the visual
part of the brain. When the subjects are shown an image of
a face, brain activity (black bars) becomes highest in the
associative and frontal regions (4, 5, and 6). Lastly, when
the subjects are retaining an image of a face in their working
memory, brain activity (red bars) is highest in the frontal
regions, while the visual areas are scarcely stimulated at
all.
It has also been observed that
distinct processes appear to be involved in the storage
and recall of items memorized with the phonological loop
and the visual/spatial sketchpad.
One thing is certain: the prefrontal cortex
plays a fundamental role in working
memory. It enables people to keep information available that
they need for their current reasoning processes. For this purpose,
the prefrontal cortex must cooperate with other parts of the cortex
from which it extracts information for brief periods. For this
information to eventually pass
into longer-term memory, the limbic system probably has to
be brought into play.
The hippocampus receives
connections from the cortex’s primary sensory areas,
unimodal associative areas (those that involve only one sensory
modality), and multimodal associative areas, as well as from
the rhinal and entorhinal cortexes. While these anterograde
connections converge at the hippocampus, other, retrograde
pathways emerge from it, returning to the primary cortexes,
where they record in the cortical synapses the associations
facilitated by the hippocampus. Thus, even in the mechanism
of memorization, we find the feedback loops so often encountered
at all levels in the living world.
For a
piece of information to be recorded in long-term memory,
it must pass through Papez’s circuit. Injuries to
this circuit can result in memory impairments.
For example, a lesion in the mammillary bodies is responsible
for an amnesic syndrome whose most classic example is Korsakoff’s
syndrome. In addition to the confabulation, confusion, and
disorientation that accompany this syndrome, patients suffer
from anterograde amnesia: they cannot store new information
in their long-term memory. The most typical cause of this syndrome
is vitamin B1 deficiency, often seen in chronic alcoholics.
LONG-TERM MEMORY
Recent research has provided a complex,
highly intricate picture of memory functions and their loci in
the brain. The
hippocampus, the temporal lobes, and the
structures of the limbic system that are connected to them
are essential for the consolidation of long-term memory.
The hippocampus facilitates associations
among various parts of the cortex, for example, between a tune
that you heard at a dinner party and the faces of the other guests
who were at the table. However, all other things being equal,
such associations would naturally fade over time, so that your
mind did not become cluttered with useless memories. What might
cause such associations to be strengthened and eventually etched
into long-term memory very often depends on “limbic” factors,
such as how
interested you were in the occasion, or what emotional charge it
may have had for you, or how gratifying you found its content.
The various structures
of the limbic system exert their influence on the hippocampus
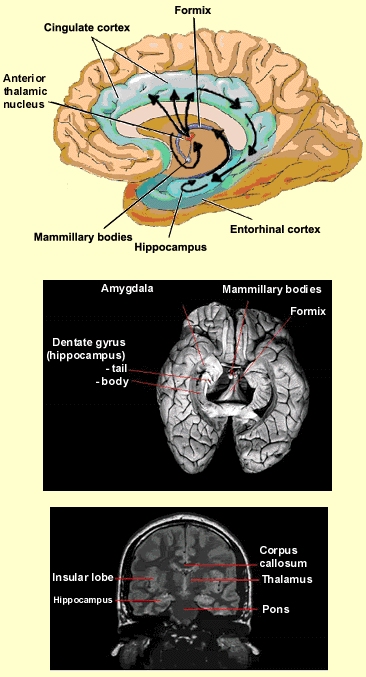
and the temporal lobe via Papez’s circuit, also known
as the hippocampal/mammillothalamic tract. This circuit
is a sub-set of the numerous connections that the limbic
structures have with one another. The diagram here shows
the route that information travels from the hippocampus
to the mammillary bodies of the hypothalamus, then on to
the anterior thalamic nucleus, the cingulate cortex, and
the entorhinal cortex, before finally returning to the
hippocampus.
Once the temporary associations of
cortical neurons generated by a particular event have made
a certain number of such “passes”
through Papez’s circuit, they will have undergone
a physical remodelling that consolidates them. Eventually,
these associations will have been strengthened so much
that they will stabilize and become independent of the
hippocampus. Bilateral lesions of the hippocampus will
prevent new long-term memories from forming, but will not
erase those that were encoded before the injury.
With this gradual disengagement
of the limbic system, the memories will no longer pass
through Papez’s circuit, but instead will be encoded
in specific areas of the cortex: the same ones where
the sensory information that created the memories was
initially received (the occipital cortex for visual memories,
the temporal cortex for auditory memories, etc.).